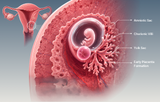
La recherche en embryologie fournit un cadre mécanistique permettant de comprendre comment un zygote totipotent unique initie des divisions cellulaires coordonnées, établit les premières identités lignagères et donne naissance aux fondements cellulaires du développement mammalien. En embryologie humaine et mammalienne, la période préimplantatoire s'étend de la fécondation jusqu'à la segmentation, la compaction, la formation de la morula, le développement du blastocyste et la préparation à l'implantation. Au cours de cette première fenêtre développementale, l'embryon subit des divisions mitotiques rapides sans augmentation globale de taille, active les programmes du génome embryonnaire, organise sa polarité cellulaire et sépare progressivement les premiers lignages extra-embryonnaires et embryonnaires. Ces processus sont essentiels non seulement pour la biologie du développement, la médecine de la reproduction et la recherche en FIV, mais également pour l'interprétation des cellules souches embryonnaires, des modèles d'embryons humains, des blastoïdes et des plateformes de biologie régénérative visant à reproduire in vitro les états précoces du développement.
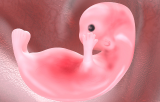
La fécondation restaure la diploïdie et initie la transition ovocyte-embryon, suivie des divisions de segmentation qui génèrent les blastomères puis, finalement, la morula. La compaction et la polarité apico-basale contribuent à l'établissement de différences spatiales entre les cellules externes et internes, favorisant ainsi la première décision de destinée cellulaire entre le trophectoderme et la masse cellulaire interne. Le blastocyste qui en résulte contient une cavité remplie de liquide, un épithélium externe de trophectoderme associé aux futurs lignages placentaires, ainsi qu'une masse cellulaire interne à l'origine des dérivés embryonnaires et endodermiques extra-embryonnaires. Les revues récentes soulignent que le développement préimplantatoire mammalien n'est pas régi par un mécanisme déterministe unique, mais résulte de l'intégration des effets de la position cellulaire, de la polarité, de la contractilité, des contacts intercellulaires, de l'hétérogénéité transcriptionnelle et des réseaux de signalisation.
Principaux marqueurs moléculaires et régulation de la pluripotence
La pluripotence dans l'embryon préimplantatoire est étroitement associée à des régulateurs transcriptionnels tels que OCT4/POU5F1, NANOG, SOX2 et d'autres facteurs associés à l'épiblaste, tandis que la spécification des lignages implique une régulation réciproque avec des facteurs associés au trophectoderme tels que CDX2. Des études d'édition génomique réalisées sur des embryons humains ont montré que l'invalidation de POU5F1 compromet le développement du blastocyste et modifie l'expression de gènes associés à l'épiblaste, notamment NANOG, ainsi que de gènes extra-embryonnaires du trophectoderme tels que CDX2, soulignant ainsi l'importance spécifique à l'espèce de OCT4 au cours de l'embryogenèse humaine. Dans les études sur le blastocyste murin, CDX2 est impliqué dans la spécification du trophectoderme ainsi que dans la répression de Oct4 et de Nanog au sein du lignage trophoblastique, illustrant la manière dont les réseaux transcriptionnels stabilisent progressivement l'identité des lignages cellulaires.
Modèles de cellules souches et systèmes de blastoïdes
Les cellules souches embryonnaires humaines et les cellules souches pluripotentes induites ont considérablement élargi l'accès à l'étude des états précoces du développement en permettant la mise en place de systèmes in vitro reproduisant certains aspects de la biologie du blastocyste. Des travaux récents sur les blastoïdes humains ont démontré que des cellules souches pluripotentes humaines naïves peuvent s'auto-organiser en structures analogues au blastocyste contenant des équivalents du trophectoderme, de l'épiblaste et de l'endoderme primitif, bien que la valeur biologique de ces modèles d'embryons humains dépende de leur capacité à reproduire fidèlement la chronologie, l'organisation spatiale et les états transcriptionnels des blastocystes naturels. Ces modèles reposent fréquemment sur des conditions de culture définies, des facteurs de croissance, des petites molécules modulant des voies de signalisation telles que FGF/ERK, TGF-β, Hippo et WNT selon les protocoles utilisés, ainsi que sur des systèmes de culture 3D, des micropuits d'hydrogel, du Matrigel ou d'autres environnements de matrice extracellulaire destinés aux essais de culture prolongée ou reproduisant des stades post-implantatoires.
Considérations expérimentales en embryologie moderne
À mesure que la recherche en embryologie évolue vers des analyses à plus haute résolution de l'émergence des lignages cellulaires, des interactions embryon-endomètre, de la pluripotence naïve et primée ainsi que des modèles embryonnaires dérivés de cellules souches, la fiabilité expérimentale dépend de plus en plus de l'utilisation de réactifs rigoureusement sélectionnés, d'anticorps validés, de consommables standardisés et d'environnements de culture reproductibles. L'embryologie mammalienne avancée nécessite ainsi non seulement une compréhension conceptuelle du développement préimplantatoire, mais également une intégration précise des outils de recherche en FIV, des marqueurs de pluripotence, des plateformes de manipulation embryonnaire et des réactifs de biologie du développement adaptés à chaque question expérimentale.