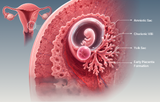
Embryology research provides a mechanistic framework for understanding how a single totipotent zygote initiates coordinated cell divisions, establishes early lineage identity, and gives rise to the cellular foundations of mammalian development. In human and mammalian embryology, the preimplantation period extends from fertilization through cleavage, compaction, morula formation, blastocyst development, and preparation for implantation. During this first developmental window, the embryo undergoes rapid mitotic divisions without overall growth, activates embryonic genome programs, organizes cell polarity, and progressively segregates the first extra-embryonic and embryonic lineages. These processes are central not only to developmental biology, reproductive medicine, and IVF research, but also to the interpretation of embryonic stem cells, human embryo models, blastoids, and regenerative biology platforms that aim to recapitulate early developmental states in vitro.
Fertilization restores diploidy and initiates the oocyte-to-embryo transition, followed by cleavage divisions that generate blastomeres and ultimately the morula. Compaction and apical–basal polarity contribute to spatial differences between outer and inner cells, supporting the first lineage decision between trophectoderm and inner cell mass. The resulting blastocyst contains a fluid-filled cavity, an outer trophectoderm epithelium associated with future placental lineages, and an inner cell mass that gives rise to embryonic and extra-embryonic endodermal derivatives. Contemporary reviews emphasize that mammalian preimplantation development is not governed by a single deterministic mechanism, but emerges from integrated effects of cell position, polarity, contractility, cell–cell contact, transcriptional heterogeneity, and signaling networks.
Key Molecular Markers and Pluripotency Regulation
Pluripotency in the preimplantation embryo is closely associated with transcriptional regulators such as OCT4/POU5F1, NANOG, SOX2, and related epiblast-associated factors, while lineage specification involves reciprocal regulation with trophectoderm-associated factors such as CDX2. Human embryo genome-editing studies have shown that disruption of POU5F1 compromises blastocyst development and alters expression of both epiblast-associated genes, including NANOG, and extra-embryonic trophectoderm genes such as CDX2, underscoring the species-specific importance of OCT4 during human embryogenesis. In mouse blastocyst studies, CDX2 is implicated in trophectoderm specification and in the repression of Oct4 and Nanog within the trophectoderm lineage, illustrating how transcriptional networks progressively stabilize lineage identity.
Stem Cell Models and Blastoid Systems
Human embryonic stem cells and induced pluripotent stem cells have expanded access to early developmental states by enabling in vitro systems that model aspects of blastocyst-stage biology. Recent work on human blastoids demonstrates that naïve human pluripotent stem cells can self-organize into blastocyst-like structures containing analogues of trophectoderm, epiblast, and primitive endoderm, although the biological value of these human embryo models depends on how faithfully they reproduce the timing, spatial organization, and transcriptional states of natural blastocysts. Such models frequently rely on defined culture conditions, growth factors, small molecules that modulate pathways such as FGF/ERK, TGF-β, Hippo, and WNT depending on the protocol, as well as 3D culture systems, hydrogel microwells, Matrigel or other extracellular matrix environments for post-implantation-like or extended culture assays.
Experimental Considerations in Modern Embryology
As embryology research moves toward higher-resolution analysis of lineage emergence, embryo–endometrium interactions, naïve and primed pluripotency, and stem cell-based embryo models, experimental reliability increasingly depends on rigorously selected reagents, validated antibodies, standardized consumables, and reproducible culture environments. Advanced mammalian embryology therefore requires not only conceptual understanding of preimplantation development, but also precise integration of IVF research tools, pluripotency markers, embryo manipulation platforms, and developmental biology reagents adapted to each experimental question.